New research using sediments from a Dutch canal has yielded important insights into the anaerobic oxidation of methane (AOM). A group of scientists from the Institute for Water and Wetland Research at Radboud University identified archaea of the order Methanosarcinales as being responsible for AOM coupled to iron and manganese reduction.
AOM proceeds as methane is oxidised with various terminal electron acceptors including sulphate, nitrite and nitrate, and the importance of oxidised metals in the process has been investigated, but the microorganisms involved have remained unknown. To address this, a culture of methanotrophs was established that was enriched with canal sediment. When this culture was supplied with nitrate as the only available electron acceptor, it became dominated by AOM-associated archaea (AAA). This culture linked methane oxidation to nitrate reduction, with N2 being the main end product, although approximately 10% of nitrate was converted to ammonium.
The enrichment culture with AAA was used for further experiments, where methane oxidation was observed following the addition of ferric citrate, nanoparticulate ferrihydrite (Fe3+) or birnessite (Mn4+). Analysis of the AAA genome showed that it could couple methane oxidation to nitrate reduction or Fe3+ reduction, or that the reverse methanogenesis pathway could also operate. The authors therefore suggest that AAA could act as a versatile methanotroph, switching electron acceptors depending on their availability.
The paper was jointly led by Katharina Ettwig and Baoli Zhu. One of the co-authors, Boran Kartaldescribed the possible application of their findings: “A bioreactor containing anaerobic methane and ammonium oxidizing microorganisms can be used to simultaneously convert ammonium, methane and oxidized nitrogen in wastewater into harmless nitrogen gas and carbon dioxide, which has much lower global warming potential.” The authors conclude that their work “may also shed light on the long-standing discussion about Fe2+ -producing processes on early Earth, when AAA-related organisms may have thrived under the methane-rich atmosphere in the ferruginous Archean oceans.”
Ettwig, K.F., Zhu, B., Speth, D., Keltjens, J.T., Jetten, M.S.M., Kartal, B. 2016. Archaea catalyse iron-dependent anaerobic oxidation of methane. PNAS, doi: 10.1073/pnas.1609534113.
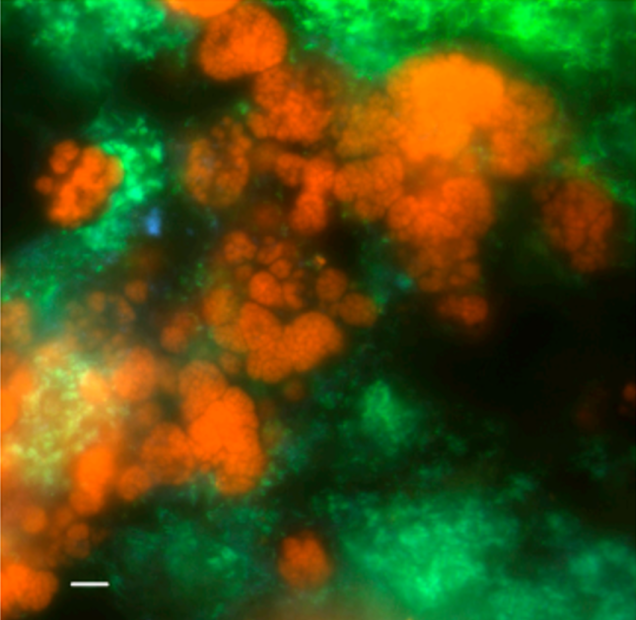
Image is from the paper and shows fluorescence in situ hybridization of biomass from the enrichment culture of AAA and M. oxyfera-like bacteria.